
间充质干细胞(MSC)在临床上的应用越来越受到重视。然而,目前临床试验的绝大多数结果都不能满足医疗需求。与胚胎干细胞以及诱导的多能干细胞和造血干细胞不同,MSC具有自己独特的功能特征。因此,与治疗相关的MSC生物学特性的一些问题需要在未来的临床应用中进行深入探讨。在这一视角下,我们重点研究了MSCs的基本和重要的生物学特性,然后分析了MSC的临床应用。我们试图为MSC治疗策略的优化提供合理的解释,以改进治疗。
01、MSC的功能特性
基于三个功能特性:体外具有多项分化潜能、免疫调节作用(低免疫原性)、促进组织器官的修复,MSC备受临床医生和科学家的关注。最近的20年涌现出了大量的基础研究和临床研究的科研成果,欧盟、日本、加拿大、新西兰、韩国、我国台湾等国家地区都批准MSC为干细胞药品上市。干细胞疗法曾被视为万能药(cure-all)[1]。理论上, 胚胎干细胞(ESC)和iPS可能是万能药,但是MSC肯定不是。然而,由于细胞生物学和传统药理学之间的差异,MSC作为药物面临着巨大的临床挑战,必须在临床上证明自己的疗效[2]。最近,一篇综述讨论了培养基、细胞来源、培养环境和存储方式的选择对MSC产品的表型和临床用途的影响,明确提出细胞质量和细胞数量是MSC临床应用的两大关键因素[3]。本文结合MSC的功能特性,深入分析MSC疗法的机理和可能影响因素,试图为MSC治疗策略的优化提供合理的解释,以改进治疗。
02、不同供体和组织来源的MSC存在变异性(异质性)
MSC存在于多种组织中,包括骨髓(BM)、脂肪(AD)、脐带血(UBC)、脐带(UC)、羊膜(AM)和牙髓(DP)。基本上,来自不同组织来源的MSCs共享相同的细胞表面标记[4]和三项分化能力[5]。虽然它们具有相同的基本功能特征,但它们之间的功能强度仍存在差异,如细胞大小、增殖潜能、分泌的细胞因子和免疫抑制。
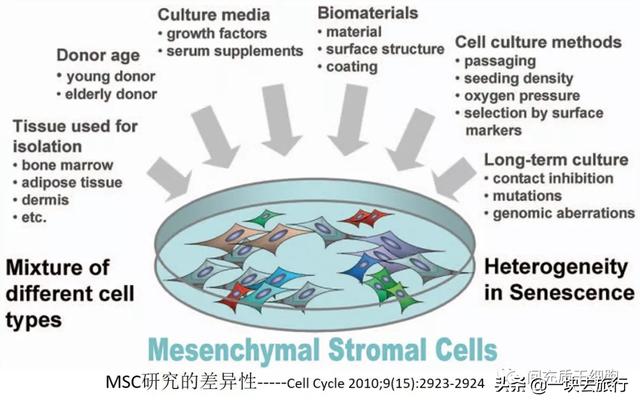
来自不同组织的MSC群体在其基因表达和分化能力方面存在巨大差异[6]。人脂肪MSC与人骨髓MSC在发育学、生物学、临床前和临床应用方面存在着一定的差异[7]。基因表达数据显示,来自脐带和羊膜的MSC具有更高的免疫调节能力,而骨髓MSC显示出更好的支持再生过程的潜力,例如神经元的分化和发育[8]。由于小鼠/大鼠的遗传背景高度一致,因此小鼠/大鼠来源的MSC的比较数据更有意义和更有说服力。与骨髓MSC相比,脂肪MSC显示出更高的增殖活性,血管内皮细胞生长因子(VEGF)和肝细胞生长因子(HGF)的产生比骨髓MSC更多,从而可以更好地治疗小鼠脑缺血模型[9]。具有高增殖率的脂肪MSC比来自相同大鼠的骨髓MSC表达更高的Nestin和神经营养因子[10]。在相同的培养体系下,马骨髓MSC的群体倍增时间(PD)在大约27个PD(第10代)后显著增加,并且骨髓MSC在第11代停止增殖,而马脐带MSC和脂肪MSC实现了大约60-80个总的群体倍增(第20-22代)[11]。马组织来源的MSC实验还表明,在相同的培养条件下,骨髓MSC比脂肪MSC和脐带MSC衰老早得多。
除了组织的多样性,利用健康人骨髓MSC开展细胞治疗之时,还需考虑到供体的变异性导致MSC在增殖动力学方面的差异[12]。来自多个捐赠者的人骨髓MSC在集落形成、细胞大小、增殖能力和免疫抑制能力方面有很大差异[13]。因为骨髓细胞的CFU-F随着年龄的增长而减少[14-16],导致无法补充祖细胞[17-21],因此CFU-F实验亦用于评估MSC质量[22]。衰老的MSC有一些变化:质量下降、分化/再生能力降低、迁移能力降低[23]。
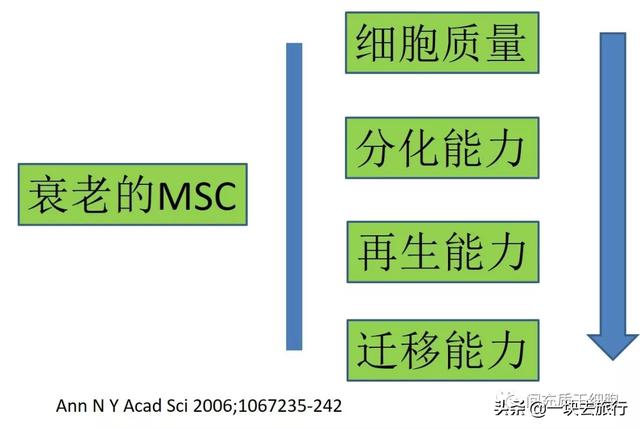
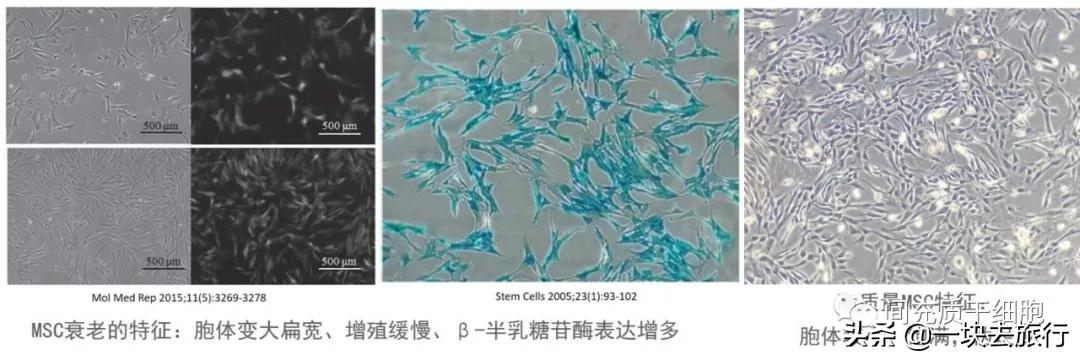
几个小组已经证明,来自老年捐赠者的MSC在人[24-26]和大鼠[27, 28]中的增殖速度都较慢。在老年供者的样本中SA-β-Gal阳性(细胞衰老的标记物)的骨髓MSC数量增加[29]。生物活性氧是正常氧代谢的天然副产品,在细胞信号转导和稳态中具有重要作用[30]。随着年龄的增长,MSC的强大抗氧化活性逐渐降低[31]。在细胞形态学方面,来自老年供体的MSC在体外培养时,MSC胞体比年轻的MSC大得多[25, 32, 33],而胞体的变大与细胞衰老密切相关[34]。年龄相关衰老的骨髓MSC降低表达细胞表面标记物CD13,CD29,CD44,CD73,CD90,CD105,CD146和CD166[35, 36]。此外,与大鼠的年轻MSC相比,机体的衰老对MSC的增殖、多潜能和代谢特征产生负面影响,老化的MSC在体外扩增过程中明显丢失了其祖细胞特征和降低了抗氧化能力[28]。有趣的是,MSC分化为脂肪细胞、成骨细胞和软骨细胞的能力是一个基本的生物学特征,不受年龄的影响[37]。此外,MSC的骨形成能力与体外分化能力无关[37-40]。
03、复制性衰老导致了大规模培养扩增的MSC的细胞质量下降
MSC的体外培养扩增不可避免地经历复制性衰老[35, 41-43],伴随着基因组不稳定性[44-47],这是MSC细胞治疗行业一个严重的障碍。MSC在长期培养过程中出现衰老,随后改变其代谢特征[28, 48],这种代谢特征的改变与线粒体融合和裂变事件相关[28, 49, 50]。复制次数的增多,常常伴随着衰老过程中的遗传损伤的积累增多[51],减弱了干细胞的可用性和功能[52]。需要注意的是,因为接种起始MSC细胞浓度不一样,即使经过相同的扩增代数,细胞的复制周期是不一致的。复制性衰老与复制周期相关,而和代数的关系没那么大。细胞核型分析显示,骨髓MSC在第18代时发生染色体异常和端粒酶缩短[53],而脐带MSC在30代之前保持染色体稳定[54]。然而,也有研究显示人类MSC似乎是遗传稳定的,在长期培养后没有显示染色体异常,并且不具有致瘤性[53, 55]。有趣的是,由天然修复蛋白质组成的血小板裂解物能支持人骨髓MSC的长期培养扩增,具有稳定染色体的作用[56]。
经过长期的细胞培养,骨髓MSC出现复制性衰老的表现是低增殖率,衰老相关的β-半乳糖苷酶出现高活性,DNA修复和抗氧化能力降低,p53和p16的表达增强[57]。p16INK4a的表达和β-半乳糖苷酶的活性与细胞面积有很强的相关性,第5代的人骨髓MSC的面积是第1代的4.8倍[58]。这些发现可用于开发基于非侵入性成像的新方法,以筛选和定量临床级细胞培养中的老化[58]。然而,另一项实验表明,p53和p16基因的表达在第15代中没有明显变化[59]。高代数的MSC也表现出p21的表达增加[60],p21是细胞周期蛋白依赖性激酶的抑制剂,并且敲除高代数MSC中的p21会增强MSC增殖能力[60, 61]。
骨髓MSC在培养到第9代时,就出现80%的SA-gal阳性细胞,即80%的MSC已经衰老了[35]。另一个实验室在培养牙髓MSC在传代10-11时出现约40%的SA-gal阳性MSC[19]。传代相关衰老的差异可能归因于不同的细胞培养系统[62, 63]。此外,高代数的MSC能触动更多的即时血液介导的炎症反应[64]并激活补体途径[64, 65],限制了它们在体内的存活和发挥功能。
衰老的MSC经历了明显的遗传和表观遗传变化[66-69]。大量证据表明miRNAs在调节干细胞功能中起重要作用[70]。干细胞的衰老也与miRNAs表达失调有关,已发现miR-335和miR-195在骨髓MSC衰老中起关键作用,改变它们的表达可以逆转MSCs的治疗效果[71, 72]。有两项研究通过微阵列或定量PCR方法证明了长期培养的骨髓MSC,会出现衰老相关miRNA表达谱的改变[35, 73]。对人脐带MSC和脐血MSC早期(P4代)和晚期(P11代)传代的microRNA图谱分析表明,来自脐带血和脐带血的MSC的衰老机制可能不同[74]。高代数(P13-P22)MSC分泌的微囊泡(MVs)(<500 nm)小于低代数(P3-P7)MSC,伴随CD105+MSC-MVs减少和miR-146a-5p增加[75]。
如上所述,长期细胞培养可能会降低MSC的治疗效果。例如,来自大鼠骨髓的低代数(P4)MSC比高代数(P40)MSC在体外诱导分化为多巴胺能样细胞的效率更高[76]。与低代数(P2-3)MSC相比,高代数(P7)MSC的条件培养基显示其神经保护功能降低[77]。使用大鼠疾病模型,发现培养扩增的小鼠MSC的心脏保护作用随着传代数量的增加而降低,并在第5代时丧失[78]。最近的研究发现,人骨髓MSC的复制性衰老是由于泛素C(UBC)表达降低所致[79]。多聚(ADP-核糖)聚合酶-1(PARP-1)通过依赖于损伤响应激酶ATM的复杂信号机制来响应DNA损伤而被激活[80]。低代数(P2-3)表达较高水平PARP-1的人骨髓MSC比高代数(P>10)的MSC对辐射或遗传毒剂诱导的DNA损伤具有更强的抵抗力[81]。PARP-1在低传代MSCs中的敲除导致对辐射诱导的凋亡的敏感化[81]。因此,在MSC长期培养中刺激UBC和PARP-1的表达是延缓复制性衰老的有效途径。虽然第3代MSC的大规模培养可以满足临床应用的需要[82],但我们也需要注意大规模培养过程中发生的细胞复制衰老[35, 66]。那么,在实际操作中,应该选取哪一代的MSC开展临床研究?显而易见的是,不同的培养工艺,MSC出现复制性衰老的代数会不一样。有些实验室培养MSC到第7代就明显出现复制性衰老,有些实验室培养到第10代才出现明显的复制性衰老。
04、不同组织来源的MSC具有不同的免疫调节能力
一些综述已经阐述了MSC和免疫细胞之间相互作用的可能机制[83-87]。MSC具有强大的免疫抑制潜力,这与剂量呈正相关,然而,工业化的大规模培养的MSC在3期试验中却未能有效治疗激素难治性急性GVHD[88]。一篇综述分析了可能原因:供体差异,表观遗传重编程,免疫原性和冷冻保存[89]。另一项关于MSC治疗激素难治性GVHD的系统综述和meta分析发现,MSC单次治疗后6个月的存活率为63%,并且与患者的年龄、MSCs培养液或提供的MSCs剂量没有差异[90]。但即使MSC具有强大的免疫调节潜能,MSC在GVHD的3期临床失败的原因仍需深入研究和调查。
长期培养是否损害了MSC的免疫调节功能?目前的研究结果并不一致。基于体外实验,MSC的免疫抑制作用从第2代到第7代没有明显区别[91]。也有研究发现人脐带MSC从第4代到第9代之间的免疫调节功能有下降的趋势[92]。有争议的是,第15代人脐带MSC的抑制活性高于第3代[93]。然而,在抗药性GVHD患者中,接受健康捐赠者低代数MSC(P1-2)的患者1年生存率为75%,而使用高代数MSC(P3-4)的患者1年生存率为仅21%[94]。长期培养(从P3到P7)对大鼠骨髓MSC的免疫豁免没有任何明显的影响[95]。
有研究提示脐带和羊膜来源的MSC的免疫调节能力明显优于骨髓和脐血来源[96]。脂肪MSC的免疫调节能力也明显优于骨髓MSC[97-99]。然而,最近的一项研究表明,脂肪MSC显示出比来自相同供体的骨髓MSC略高的免疫抑制能力,但是没有统计学差异[100]。这一结果与小鼠实验一致,即来源于同一个小鼠的脂肪MSC和骨髓MSCs在抑制T细胞增殖的免疫抑制能力上没有差异[101]。有趣的是,MSC的免疫调节能力和自身的增殖潜力之间没有明显的关系[13, 102]。因此,我们需要明确影响MSC免疫抑制能力的因素,然后选择免疫抑制能力最强的MSC用于免疫性疾病的治疗。
05、症环境可能不利于MSC发挥治疗作用
与传统的化学药物不同,MSC是一种与环境相互作用的活细胞。在某些条件下,MSC可能作为促炎细胞,并交叉呈现可溶性外源抗原作为其抗原呈递细胞特性的一部分[85, 103]。干扰素-γ刺激骨髓MSC上调MHC-II分子的表达,并将外源性抗原呈递给T细胞[96, 104]。有趣的是,炎症环境并没有增加来自脐带、脐带血和羊膜的MSC的HLA-DR表达[8, 96, 105-107]。动物实验表明,干扰素-γ与T细胞和巨噬细胞分泌的TNF-α联合诱导骨髓MSC凋亡,从而抑制骨髓MSC对骨愈合的影响[108, 109]。炎性因子IL-2激活的NK细胞和CD3/CD28激活的T细胞均可通过Fas/FasL途径诱导MSC凋亡[110, 111]。炎症环境也可以增加HLA-DR的表达,从而加速MSC的清除[96]。炎性细胞因子也通过诱导BECN1的表达协同诱导MSC自噬,这削弱了MSC治疗实验性自身免疫性脑脊髓炎的疗效[112]。有争议的是,在用干扰素-γ[113, 114]或白细胞介素-17[115]预处理后,骨髓MSC的治疗效果增强。高水平的血清干扰素-γ可以预测活动期类风湿关节炎患者对骨髓MSC治疗的良好临床反应[116]。与正常脊髓组织提取物相比,慢性损伤脊髓组织提取物显著促进MSC分泌VEGF[117]。从创伤脑组织和缺血脑组织制备的提取物也可以促进MSC中各种神经营养生长因子的表达[118, 119]。
急性肝功能衰竭的猪实验发现,严重的炎症环境限制了MSC的效果,因为MSC在严重炎症的肝脏环境中存活率低[120]。MSC未能提高伴有全身性炎症的慢加急性肝功能衰竭患者的存活率,但是减轻炎症明显有利于提高MSC的治疗效果[121]。在一项MSC治疗儿童激素难治性GVHD的多中心临床研究中,MSC在疾病早期介入治疗比晚期治疗更有效[122]。这些数据清楚地表明,严重的炎症环境可能会减弱MSC的治疗效果。重要的是,同种异体MSC输入可以诱导CD4+和CD8+T细胞的记忆表型[123],这表明当相同来源的同种异体MSC重新输注到患者体内时,会发生更快的清除。如上所述,输注的MSCs最终被宿主T细胞、NK细胞和巨噬细胞清除或杀死。因此,需要多次输注骨髓间充质干细胞,以确保预期的显着治疗效果。
06、MSC输入后的体内分布
曾经期待MSC输注后可以像造血干细胞一样在体内长期存活,伴随终身。然而,大量的研究表明输入的MSC不能在活体中长时间存活[123-126]。肺脏是静脉注射MSC需要跨越的首个屏障,因为由于肺血管系统的特性,大多数静脉输入的MSC被困在肺内[127-129]。MSC静脉输注后1小时,约50-60%的MSC保留在肺内,3小时后比例降至30%并维持96小时[130, 131]。在存在肺损伤的情况下,被困在肺中的MSC数量增加[124, 132]。当MSC滞留在肺脏时,MSC被局部微环境激活,分泌大量的抗炎因子TSG-6,这有利于减少炎症和减少心肌梗死面积[132]。穿过肺脏后,MSC到达肝脏、肾脏和脾脏[26, 126, 127, 133-135]。狒狒实验证据表明,经外周静脉输入骨髓MSC,自体MSC和同种异体MSC的体内分布没有明显差异[125]。与同基因或免疫缺陷小鼠相比,健康免疫完全异基因小鼠的MSC存活时间明显缩短[123]。动物实验表明,免疫系统负责清除输入的MSC。MSC几乎不表达MHC II类分子(HLA-DR),这个MHC II类分子主要是引起免疫反应,为何MSC离开机体内原来存在位置的微环境后,终究会被机体免疫细胞清除掉?不管是血管内注射还是局部注射,MSC都不能长期存活。
已证明MSC表达多种细胞表面粘附分子[136, 137]和参与MSC迁移和粘附的蛋白酶[138, 139]。与骨髓MSC相比,脐血MSC更容易通过肺部,因为脐血MSC胞体更小,并且表达的CD49f和CD49d水平明显高于骨髓MSC[133]。有趣的是,用硝普钠预处理后,肺微血管系统中的MSC截留明显减少[126, 128]。这种现象归因于硝普钠产生的一氧化氮(NO)引起的血管扩张,增加了血液流量,随后导致MSC快速通过肺部。因此,硝普钠预处理有效增强了MSC修复CCl4诱导的小鼠肝纤维化的治疗效果[140]。在一项临床试验中,用111In-oxine(放射性示踪元素)标记的自体骨髓MSC通过外周静脉注入失代偿期肝硬化患者,标记的MSC首先在肺中积累,然后在接下来的几小时到几天内逐渐转移到肝脏和脾脏[141]。
动物实验也证明,直接注射到组织中的MSC在体内同样不能维持长期存活,如脑[142, 143],关节腔[144, 145],心肌[146, 147]。颈动脉注射MSC也存在同样的现象[148-150]。由于操作简单,MSC颈动脉注射被用于治疗脑部疾病,但是需要注意形成细胞团后导致的微小血管的栓塞。局部微量注射到大鼠脑纹状体的骨髓MSC可以沿着已知的途径迁移到胼胝体和大脑皮层,迁移途径类似神经干细胞迁移到大脑的连续层[142]和脑血管[143]。通过静脉注射的人MSC也可以跨越血脑屏障迁移到创伤性脑损伤大鼠的受损皮质边缘[151],可能是因为脑外伤后血脑屏障的通透性增加[152]。出乎意料的是,人骨髓MSC和大鼠骨髓MSC通过大鼠的颈内动脉输送后,在脾脏、肾脏、肝脏中同时发现了人MSC和大鼠MSC的存在,这说明MSC经过颈内动脉后迁移到内脏组织[149, 150]。
07、优化MSC治疗方案
治疗策略应包括足够剂量的MSC、治疗途径、治疗时机,和/或与其他药物的联合。MSC的质量与不同的培养体系和来源密切有关,甚至直接影响到MSC的动物实验和临床研究结果[63]。MSC的临床前研究数据并不能很好地指导临床应用的方案确定。在临床研究中,每个患者使用的MSC数量从4000个到数亿个不等。局部介入治疗的最低剂量发生在用4500个MSC治疗的股骨头坏死的临床病例[153, 154]。糖尿病肢体大疱性疾病的介入治疗最高剂量为8.6亿个MSC[155]。全身输液量一般为(1-10)x106/kg。目前临床静脉输入的最高剂量出现在MSC与造血干细胞共同移植时,剂量为10x106/kg[156]。有趣的是,一项meta分析表明,剂量并不影响接受单次输注MSC治疗的急性GVHD患者的存活率[90]。理论上,MSC的最佳剂量取决于不同的疾病和移植途径,可能存在一个范围,在这个范围内,剂量越高,疗效越好。
MSC在分布和代谢上与传统药物有很大不同,在健康和病体中的分布也是不同的,因为MSC能够主动趋化到损伤部位。在动物实验中,传统药物需要多次给药以维持稳定的血药浓度,而现在尚未意识到MSC也需要多次注射来维持一定的有效细胞浓度。人们期望MSC只用一次移植就能治愈一些疾病,就像造血干细胞一样。事实上,研究表明,用于治疗心脏病的MSC治疗,只有不到1%的移植细胞在注射后1周内存活[157, 158],这意味着需要多次输注来维持治疗效果。如果进行一次治疗,低剂量和高剂量MSC之间的疗效没有差异[159]。反复输注MSC导致肝衰竭疾病更好的临床结果[160, 161],而单一的MSC治疗未能产生明显的长期(48周)疗效[162]。值得注意的是,当MSC治疗与药物[159]和其他治疗[163, 164]相结合时,将获得良好的临床结果。此外,尽管新鲜和冷冻的骨髓MSC都具有相同的生长特征,但来自新鲜骨髓MSC具有更高的活力[165]。
此外,在使用MSC治疗不同疾病时,需要更多的证据来确定最佳剂量。我们先前已经讨论过MSC治疗GVHD的策略优化[166]。即使患者具有相同的体重,在治疗免疫性疾病和其他疾病时,MSC的剂量也可能不同。因此,MSC的剂量与疾病的亚型密切相关。例如,IV型GVHD可能需要比II型更高剂量的MSC。这一假设需要在未来的临床研究中得到证实。
08、分化可能不是MSC的治疗机制
干细胞生态位(龛,Niche)是指与干细胞相互作用以调节细胞命运的微环境,相当于干细胞的窝,例如骨髓中的造血干细胞生态位[167-170],毛囊中的毛囊干细胞生态位[171-173],以及隐窝碱基中的肠道干细胞生态位[174-176]。这个干细胞生态位的主要功能是锚定和滋养干细胞,既避免干细胞的快速耗竭,同时避免过度旺盛的干细胞增殖[177]。一般情况下,干细胞在其生态位中保持静止状态。当接收到刺激信号时,周围的微环境促进干细胞自我更新或分化成子细胞以形成新的组织[178]。如果患者的自体造血干细胞被药物或辐射杀死,腾出空间来接纳移植进来的造血干细胞,在这个生态位里面,新移植进来的造血干细胞才能发挥再生整个血液和免疫系统的功能[179]。
MSCs已被证明在体内[180, 181]和体外[182]可分化为成骨细胞。在啮齿动物成骨发育不全(OI)模型中,在骨髓MSC移植之前对骨进行照射处理,这有助于骨髓MSC归巢到骨质生态位中,然后向成骨分化[183, 184]。干细胞生态位提供干细胞的稳态微环境,控制干细胞的增殖活动,维持干细胞群体。但身体内的MSC生态位在哪里?有研究提示MSC生态位类位于血管周围,并与血管密度相关[185-187]。骨髓MSC在体内的动态分布表明,如果受者的骨髓未经药物或辐射处理,则只有约0.4%的注入MSC能够迁移到骨髓中[126, 133]。因此,可能MSC只有迁移到骨髓的血管周围空间时(MSC生态位)才能避免机体免疫系统对MSC的清除,从而有机会向成骨分化。这也解释了为何不管是血管内注射还是局部注射,MSC都不能长期存活。
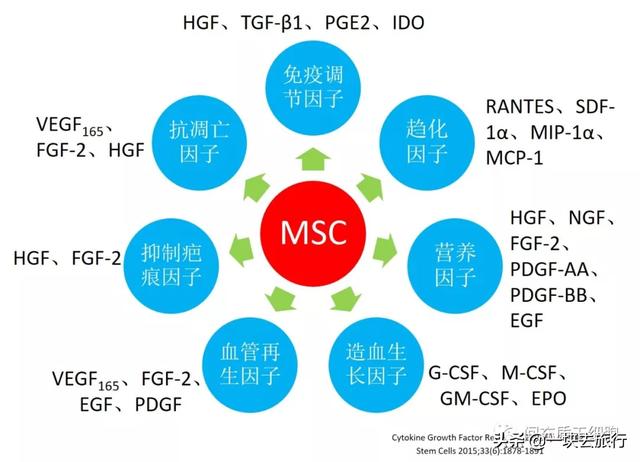
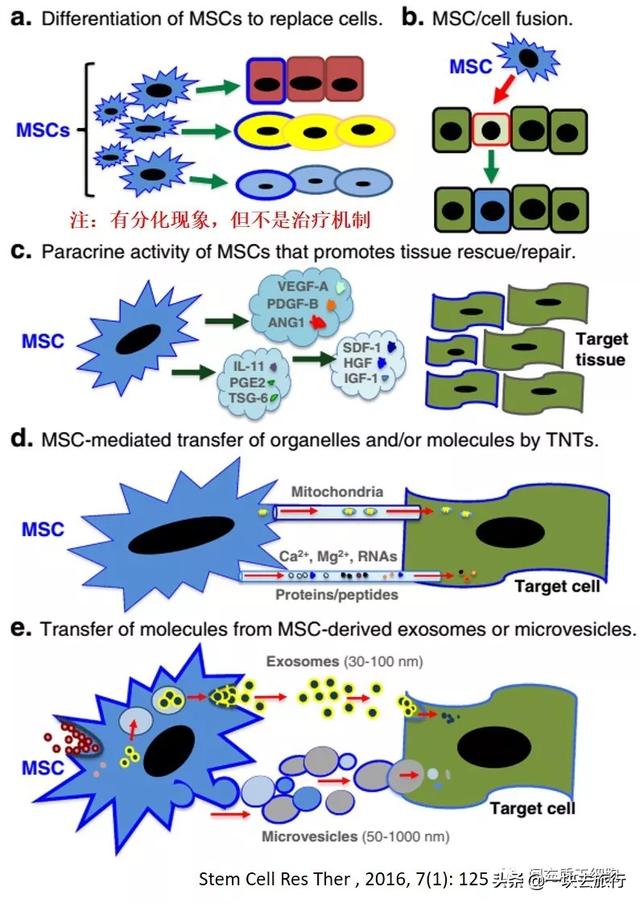
绝大多数输入的MSC并不能长期存活,这也是MSC安全性的一个重要原因,而且MSC的治疗机制是由于分泌的可溶性因子[13, 188-192],例如改善心力衰竭的VEGF[193],HGF用于多发性硬化症[194]和肝脏疾病[195, 196],IGF-1和EGF用于伤口修复[197]。这些因子通过减少炎症和减少组织细胞的凋亡或通过刺激组织内源性干细胞的增殖和分化来增强受损组织的修复[198-201]。此外,MSC通过分泌营养分子来支持造血,包括细胞因子和生长因子[202-204],这已被动物模型实验所证实[198, 205-207]。基于体内动态分布,MSC更有可能通过“触摸即走(touch and go)”[83]或“击中即跑(hit and run)”[191]机制发挥其治疗作用,在迁移到受损器官后,MSC分泌应激诱导的治疗分子或直接与靶细胞相互作用,然后被机体清除。
09、健康人和疾病患者的MSC
基本上,所有含有结缔组织的器官都含有MSC[208]。MSC具有强大的免疫抑制能力,为什么自身免疫性疾病仍然会发生在人类身上?不断有证据表明,从自身免疫性疾病患者分离出来的MSC具有形态和一些功能的异常[209-215]。来自再生障碍性贫血[214],多发性骨髓瘤患者[213],系统性红斑狼疮[209],类风湿性关节炎[215]和特发性血小板减少性紫癜(ITP)患者[212]的骨髓MSC的免疫抑制功能有不同程度的受损。与健康人真皮来源的MSC相比,银屑病患者真皮MSC在P3和P5代表现出异常的与炎症和基本细胞活性相关的基因表达模式[216]。转录组分析揭示了来自多发性骨髓瘤患者的骨髓MSC在细胞周期、免疫调节功能和成骨细胞分化方面有异常改变[217]。此外,自体骨髓MSC输入治疗未能改善系统性红斑狼疮的疾病活动进展[210]。那么自身免疫性疾病的发生和MSC的免疫抑制功能异常有没有关联,谁是因,谁是果?可能是MSC免疫抑制能力受损未能抑制自身免疫性疾病的发生和发展。
此外,股骨头坏死患者骨髓MSC的增殖和成骨分化能力降低[218-220],这可能与自体骨髓中MSC的异常lncRNA表达谱[218]和FZD1基因的异常CpG岛甲基化[221]有关。虽然骨髓MSC在临床相关剂量下对化疗药物的杀伤具有一定的抵抗性[222, 223],但大剂量化疗药物会对骨髓基质造成严重损害,从而导致造血支持能力显著降低[224-228]。
010、总结和展望
自1995年首次临床应用以来[229],MSC已被用于治疗多种疾病。然而,MSC的疗效并不令人满意,特别是在大规模的3期临床试验中未能达到预期的效果。在查阅大量MSC细胞生物学和临床研究文献的基础上,总结出影响MSC疗效的三个重要因素,即MSC的细胞质量、治疗策略和疾病适应证的选择。
如上所述,来自不同组织来源的MSC具有一定的功能差异,主要表现在增殖能力、分泌的细胞因子谱、衰老表型和免疫调节方面,这可能与最佳治疗效果密切相关。但是如何定义MSC的质量呢?这是一个非常重要的问题,同时也很难澄清。也许“生物学效力”(Bio-Efficiency)这个术语能很好的描述MSC的细胞质量。生物学效力广泛应用于医学药理学和疫苗等领域。MSC有两个惊人的功能:对不同免疫细胞的免疫调节和促进组织再生的能力。MSC的免疫调节机制与修复组织的机制明显不同。因此,生物学效力必须与特定的疾病相联系。
生产培养工艺上的差异导致了MSC功能特征的变异性,包括其最先被证明的成骨分化和支持造血的能力[230],这表明需要优化标准化制造程序[231]。更重要的是,为了在未来的大规模临床试验中获得令人满意的结果,研究者需要关注培养MSC的质量和治疗策略,而且不同的疾病,其对应的治疗策略也应该不同。而MSC供应商则需要关注整个MSC生产制备工艺,在“质量源于设计(QbD)”的指导原则下,提供稳定的高品质的MSC产品。
参考文献:暂不提供

